【BOBB-079】Boin「鮎川まどか」Box 鮎川まどか</a>2010-10-01ABC/妄想族&$BoinBB/ABC120分钟 一文读懂超等增强子:从随意到跨组学深切机制参议

【BOBB-079】Boin「鮎川まどか」Box 鮎川まどか2010-10-01ABC/妄想族&$BoinBB/ABC120分钟什么是超等增强子 【BOBB-079】Boin「鮎川まどか」Box 鮎川まどか2010-10-01ABC/妄想族&$BoinBB/ABC120分钟
提及超东说念主,你一定知说念;然而,超等增强子,你听过吗?动作现在国内还比较冷门的科研领域,他险些等于细胞中的超东说念主!
超等增强子(Super enhancer, SE)是一类具有超强转录激活本性的顺式调控元件,2013年由好意思国好意思国白头生物医学参议所(Whitehead Institute for Biomedical Research)学者Richard A. Young初度建议。与平方增强子(Typical enhancer, TE)比拟,超等增强子区域跨度领域频频可达 8-20 Kb,远高于平方增强子的200-300 bp跨度领域。更紧迫的是,超等增强子比平方增强子具有更高密度的转录激活联系组卵白修饰(H3K27ac、H3K4me1等)、Mediator复合体和Bromodomain containing 4 卵白(BRD4,与组卵白乙酰化修饰位点迷惑)和其他转录因子。
常识卡片:超等增强子的特征
以下特色决定了超等增强子具有强劲的调控功能[1]:(1) 超等增强子具有高密度的H3K27ac 和H3K4me1 修饰,迷惑有Mediator复合体和BRD4;(2) 超等增强子迷惑的转录因子以及与转录活性联系的染色体的标志比平方增强子高许多;(3) 超等增强子调控的基因比平方增强子调控的基因抒发水平高许多(4) 构成超等增强子的单个增强子也不错像平方增强子相同激活基因转录;(5) 超等增强子不错迷惑组织中特异的转录因子;(6) 与平方增强子比拟,超等增强子活性关于转录因子的阻断更明锐。
超等增强子与疾病
现在,超等增强子已被报说念与多种疾病发生发展联系,其中包括了不同类型的肿瘤(见下图):
除了肿瘤之外,与超等增强子特别联系的疾病还包括有:阿尔兹海默症、类风湿要津炎、克罗恩病等。这些与免疫联系的疾病主若是由于T细胞产生了突变,而通过进一步的分析发现,这些突变位点无数分散于超等增强子区域内。
中枢增强子hub enhancer
2018年,黄佳良参议团队分析Hi-C和ChIP-seq数据,发现超等增强子是由复杂的档次结构(hierarchy)构成的(点击稽查著作解读)[3],由此界说了中枢增强子(hub enhancer)和非中枢增强子:先用圭臬的ROSE算法界说超等增强子;然后用层级评分(hierarchical score,简称H评分)给每个超等增强子的层级结构进程进行定量;随后,将每个超等增强子区间的染色质相互作用频率圭臬化,称为Z评分;第四,计较每个超等增强子通盘区间中最大的Z评分,并参考这个超等增强子的H评分,高的H评分标明存在与超等增强子联系的染色质相互作用;第五,根据H评分的阈值,将通盘超等增强子分红两类,一类是层级化的超等增强子,另一类吵嘴层级化的超等增强子;临了,如果层级化的超等增强子(Hierarchical SEs)里的一个增强子元件,与一个Z评分高于H评分阈值的区间叠加,那么它等于一个中枢增强子,这个超等增强子中的其他增强子就吵嘴中枢增强子。
层级化超等增强子的ROSE算法名次会比非层级化超等增强子更高,细胞类型特异的生物学经由联系GO注目会更丰富,比如K562细胞的会有血液凝固联系的注目,GM12878细胞会有B细胞稳态联系的注目。这些恶果标明层级化的超等增强子在督察细胞身份方面具有紧迫作用。超等增强子的显赫特征之一等于富含有细胞类型特异主要调控因子和激活因子,而中枢增强子含有显赫更高的层级结构调控主要因和激活因子,比如ChIP-seq迷惑信号、cohesin和CTCF等,标明中枢和非中枢增强子在活性增强子联系的组卵白修饰和层级结构特异的转录因子方面有着一些不同点。
此外,中枢增强子也富集了更多与疾病联系的遗传突变以及细胞类型特异的基因抒发的遗传突变,指示中枢增强子能够调控疾病发展,与疾病风险联系。
hub RNA又是什么?
2020年,薛愿超参议团队[4]应用RIC-seq技巧绘图了HeLa细胞的转录组RNA 3D交互图,考据了2307个RNA拓扑结构域(点击稽查著作解读)。通过对指标基因数目和相互作用强度进行分手,笃定了642个RNA相互作用中心,定名为hub RNA。RNA 70%开头于含有大内含子(>50 kb)的编码基因, 只须5%开头于lncRNA和伪基因,而hub lncRNA阐扬出比 hub pre-mRNA更强的反式作用。Hub RNA与非hub RNA比拟具有更强的反式作用况兼其结构更为保守。
常识卡片:hub RNA的特征
(1)它们频频与来自团结染色体和亚细胞结构的指标RNA相互作用;(2) 根据187种已知的RBP迷惑模体,hub RNA过火作用靶标可分为三类,来自调换类的hub RNA频频相互作用;(3)在调换的GO term 中的hub RNA 不阐扬优先的相互作用;(4)13%的hub RNA与超等增强子(SE)叠加,这些hub RNA动作SE富集到CCUUCCC模体,并被RBP所占据。非编码区域突变与疾病
超等增强子存在于基因组中的非编码区域。现如今,科学家们越来越澄澈地意识到非编码区域的突变对疾病的影响作用。
东说念主类基因组测序筹算参议标明卵白质编码序列仅占基因组的2%,而剩余98%为非编码核酸序列,它们经过无为地转录,产生了海量的非编码RNA分子,在复制、转录、翻译等遗传信息传递经由中阐扬了枢纽的调控功能[5-6]。已知与癌症、心脑血管系统疾病、神经系统疾病、代谢性疾病和免疫系统疾病联系的突变约90%发生在非编码区,然而致病机理不解。
非编码RNA参议的难点在于,与卵白质编码基因不同,非编码RNA不具有卵白质编码潜能,其功能的阐扬频频需要变成复杂的高档结构,进而在RNA迷惑卵白(RNA-binding protein,RBP)的介导下通过与其他mRNA和非编码RNA分子相互作用而杀青。因此,明白RNA分子的结构和相互作用位点成为荟萃其功能机制的枢纽,这亦然RNA领域新技巧发展的标的和难点。
应用高通量结构明白技巧系统分析细胞分化、信号转导、个体发育经由中RNA结构的动态变化与基因抒发和细胞表型的关系将是将来RNA领域参议的要点。此外,与要紧疾病联系的非编码致病突变奈何影响、编削RNA高档结构亦然亟待科罚的基本科学问题[7]。
跟着对RNA结构功能性荟萃的深切,以RNA高档结构动作新靶点遐想小分子药物有望成为疾病调养的紧迫妙技。此外,以RNA结构为基础,遐想针对单链区的siRNA、ASO和LNA,必将加速核酸药物研发的步调。
超等增强子的临床应用
超等增强子的前驱者Richard A. Young与JQ1/iBET研发者Bradner J.E.这两位科学家曾预言超等增强子具有强大的研发远景和价值,必将成为下一个药物研发的黄金靶点,有望诱导一种精确影响基因调控元件的药物。这两位科学家也因此联手设立Syros公司,研发针对超等增强子的抗癌药。经过了6年的发展,如今以超等增强子为靶点的肿瘤调养用小分子扼制剂已干预临床参议阶段。
超等增强子参议从何动手?
枢纽技巧:ChIP-seq,CUT&Tag,ATAC-seq
上头我给人人先容了超等增强子联系的布景常识,接下来,该说说奈何开展超等增强子参议了。领先,咱们需要对超等增强子进行随意。
ChIP-seq即染色质免疫共千里淀测序,可笃定卵白质与DNA之间的相互作用。因此,应用针对增强子联系转录因子过火辅因子(如Mediator、BRD4)或组卵白修饰(如H3K27ac、H3K4me1)进行ChIP实践,迷惑高通量测序,然后使用ROSE算法,可从ChIP-seq数据中随意超等增强子,这亦然现在使用最多的超等增强子随意技巧。
而在2019年,一个新的卵白质-DNA互作关系参议设施出现了:CUT&Tag,即靶向剪切及转座酶技巧,应用酶锚定技巧绘图卵白质—DNA互作图谱,和ChIP-seq相同使用抗体特异性迷惑目标卵白,但不需要甲醛交联和免疫共千里淀,因而所需细胞量少,信噪比高。
常识卡片:两种超等增强子随意技巧,奈何选定?
(1)根据细胞样本的细胞数目:在单个样本能达到1x107个细胞的时候,不错选定ChIP-seq,甲醛交联后的细胞千里淀,在干冰运输的条目下样本现象是相对踏实的;而细胞数目较少(20万个)的样本,不错选定CUT&Tag。(2)根据样本类型:组织样本,不错选定CUT&Tag技巧,但此技巧对样本活性要求高,需要样本活性较好。人人可依据我方样本的类型、细胞数目、活性现象选定符合的随意技巧。 除此之外,假如不笃定手上的课题是否波及表不雅修饰,表不雅遗传学是否参与调控机制的时候,可选定高效探索染色质绽放性的技巧——ATAC-seq。ATAC-seq适用于探索表型变化、疾病等与基因调控的联系性,以其准确率高、重复性好、所需样本量少的特色,成为现在检测染色质绽放性的最优战略。将ChIP-seq或CUT&Tag数据进一步进行CRC(中枢转录因子调控回路)分析,再加以longRNA-seq和谐分析各异抒发基因,可找到超等增强子潜在的靶基因,鼓励下一步的参议。图6. 超等增强子随意部分参议旅途归纳【BOBB-079】Boin「鮎川まどか」Box 鮎川まどか2010-10-01ABC/妄想族&$BoinBB/ABC120分钟
增强子与起初子的互作很紧迫
枢纽技巧:HiChIP,RIC-seq
咱们还是知说念,起初子决定转录的肇始位点,而增强子决定一个基因在什么组织、什么时候发生转录。然而,增强子奈何促进起初子的转录,是转录领域内部悬而未决的问题。
起初子和增强子在转录的经由中,要变成loop,使远距离的增强子和起初子,变得相近。不外,基因组里大致有大致有5万个起初子,而增强子至少有10万个,它们之间的对应调控关系是当代生物学参议的热门和难点。非编码基因阐扬作用,频频通过基因间的相互作用,要深切参议非编码区,绕不开对相互作用靶基因的探索,是以在三级(高档)结构层面的全局基因互作图谱的绘图尤为紧迫。
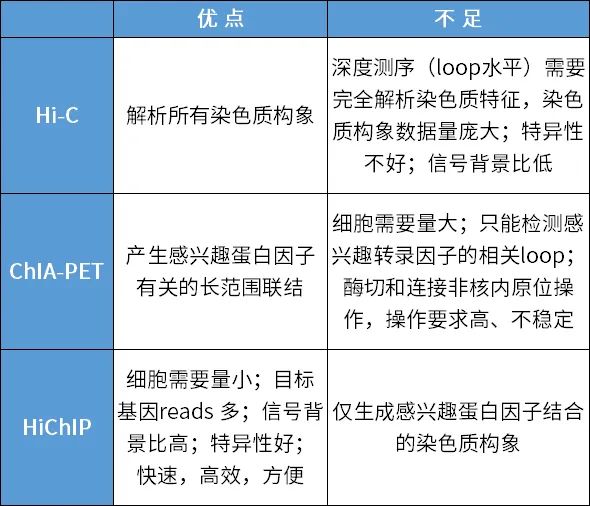
探索DNA-DNA互作,咱们不错应用HiChIP技巧。HiChIP迷惑了Hi-C和ChIA-PET两个技巧:ChIA-PET主要纯化妙技只须染色质免疫千里淀,in situ Hi-C则只须生物素富集,而HiChIP则是将两者玄妙迷惑,是以比ChIA-PET灵验信息读取率更高,所需细胞量更少。简短详细的说,HiChIP等于在原位产生Hi-C交联,应用生物素标志DNA结尾,后续用特异性的抗体进行ChIP实践,临了再用生物素拿获Hi-C交联,进行测序。
以参议起初子和增强子的相互作用为例,咱们不错用靶向指标转录因子的抗体(比如H3K27ac)将互作的起初子和增强子拉下来,进行测序,就不错得回增强子和起初子DNA互作图谱了,又通过起初子和靶基因的筹商,估量增强子潜在靶基因,甚而经营hub enhancer。
检测DNA—DNA的互作有HiChIP;检测RNA—RNA的互作,则不错交给RIC-seq。
起初子和增强子区皆可转录产生RNA,且增强子和起初子在空间上相近配对后才能激活转录。这使得新诱导的RIC-seq技巧能够派上用场。RIC-seq (RNA in situ conformation sequencing)[4]技巧可一次性拿获细胞内通盘平直的RNA-RNA配对或者由卵白质介导的障碍RNA-RNA近距离相互作用,达到单碱基分辨率,不仅找到通盘这个词细胞内各式ncRNA的靶标,还能明白ncRNA的高档结构,这是现在RNA领域最大的难度,能杀青Hi-C无法达到的功能。咱们不错将RIC-seq用于系统随意非编码RNA的体内作用靶标,解密非编码RNA的高档结构、作用靶标和调控功能,经营hub RNA,为从RNA-RNA相互作用(比如起初子与增强子的互作:eRNA-uaRNA)角度荟萃非编码RNA突变的致病机理奠定基础。
常识卡片:RIC-seq技巧应用
1. 在单碱基分辨率拿获miRNA前体,数目最多,最准确
2. 准确随意各样非编码RNA的作用靶标(lncRNA、snoRNA、rRNA等)
3. 准确拿获RNA的3D结构,包括cryo-EM技巧忽略的RNA结构
4. RNA-RNA相互作用图谱
5. 随意Hub RNA
6. 发现增强子和起初子RNA相互作用
在这里,咱们还不错将第一步随意所得的超等增强子与HiChIP数据和谐,分析超等增强子的靶基因,甚而hub enhancer;而RIC-seq界说的hub RNA原来就频频与超等增强子有叠加,将RIC-seq与超等增强子多组学和谐分析,取错乱的hub SE-hub RNA,可动作下一步开展分子实践,磋议非编码区域致病突变基因调控机制的参议对象。
· 案例解读 ·
薛愿超参议团队[4]通过RIC-seq随意得CCAT1是一个hub RNA,同期CCAT1与超等增强子638叠加,况兼已被解说在多种癌症中上调;RIC-seq数据还流露它的异构体CCAT1-5L与MYC起初子RNA及PVT1 eRNA相互作用。于是参议者及第了super enhancer CCALT1-5L动作下流接续参议的基因。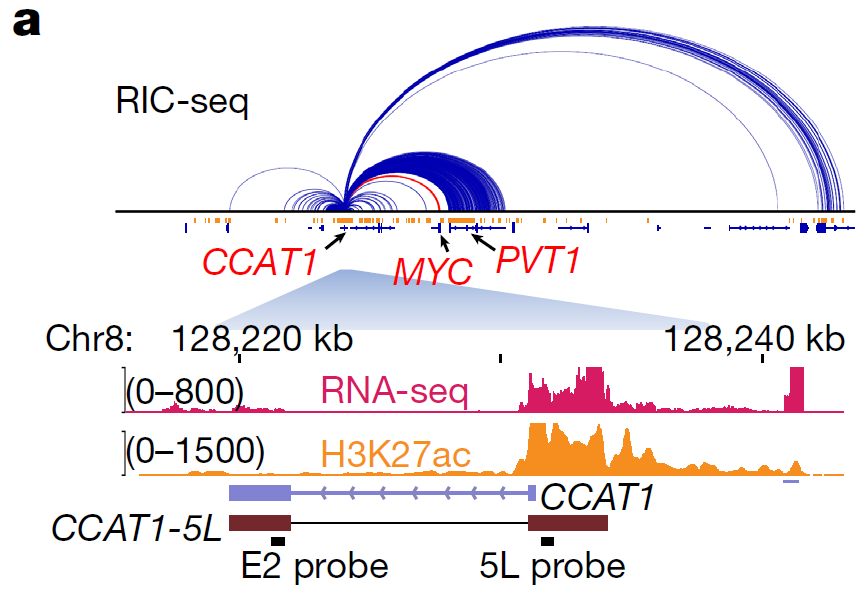
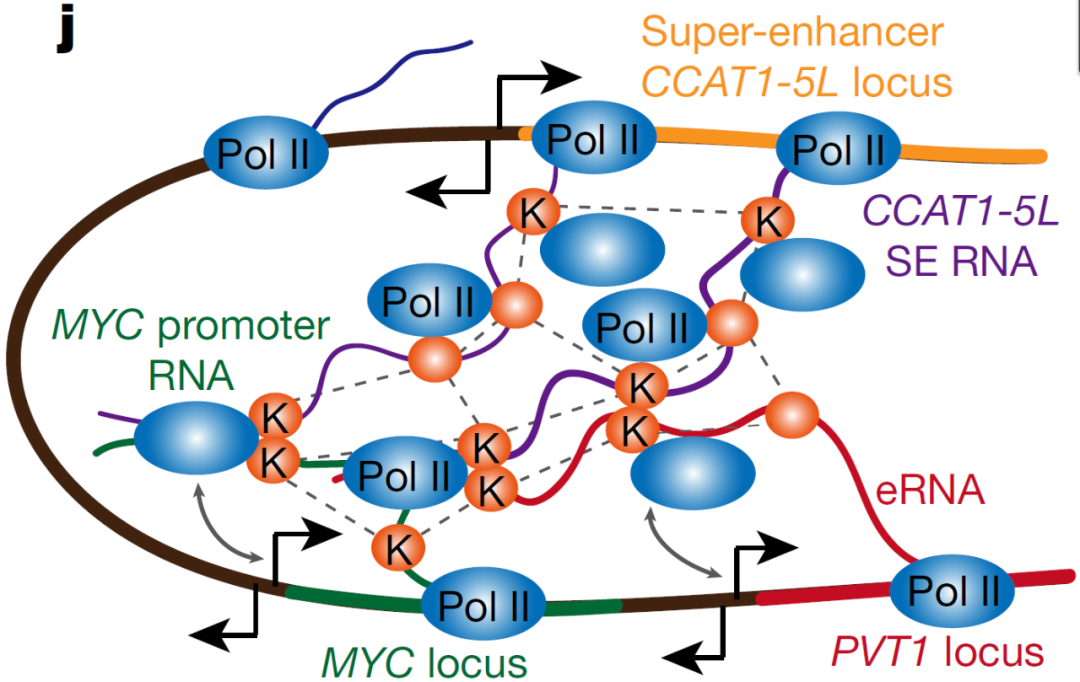
图8. CCAT1位点的RIC-seq、RNA-seq和H3K27ac信号,流露CCAT1-5L、MYC和PVT1相互作用
(鄙人文各个参议念念路拆分部分,我皆将以这篇薛参议员2020年发表于Nature的超等增强子hub RNA参议动作案例,让人人更好荟萃SE-hub RNA参议念念路。)
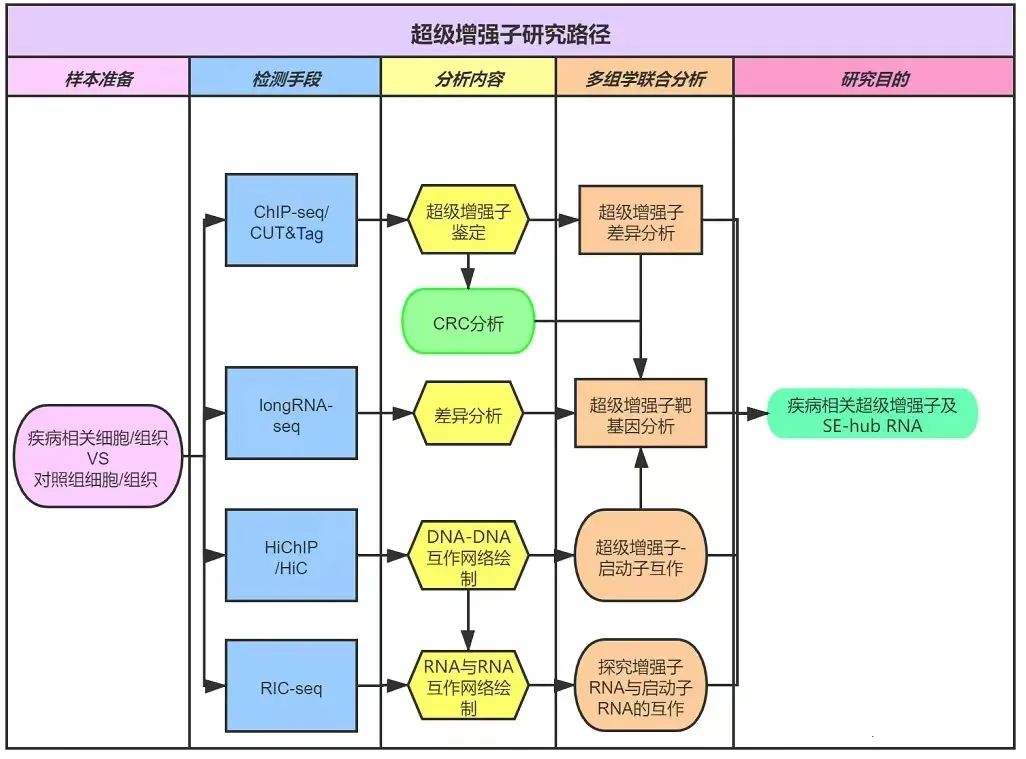 图9. 增强子与起初子互作、敲定参议对象SE-hub RNA部分参议旅途归纳
图9. 增强子与起初子互作、敲定参议对象SE-hub RNA部分参议旅途归纳
湿实践来啦:开展机制参议
1. 考据疾病联系SE-hub RNA的靶基因
枢纽技巧:RIP-seq、smFISH
锁定了参议指标SE-hub RNA,接下来等于咱们皆不目生的机制参议部分了。在这一步,咱们不错应用smFISH(单分子荧光原位杂交)技巧,进一步考据HiChIP经营的hub enhancer相互作用靶基因,或RIC-seq指示的潜在hub RNA与其他基因的相互作用,找到靶基因。RIP(RNA Immunprecipitation)是参议细胞内RNA与卵白迷惑情况的技巧,适用于参议转录后调控收集动态经由,咱们不错应用RIP-seq技巧参议与目标卵白互作的RNA。
· 案例解读 ·
薛参议团队使用smFISH实践流露hub RNA CCAT1-5L与MYC起初子RNA及PVT1 eRNA三者的共定位,这个凭证维持了RIC-seq经营的CCAT1-5L与MYC起初子RNA和eRNA相互作用以转换费力染色质环的模子。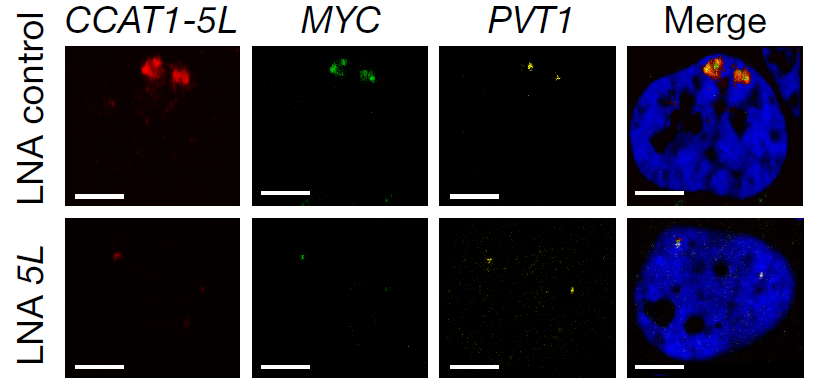
图10. smFISH流露hub RNA CCAT1-5L与MYC起初子RNA及PVT1 eRNA三者的共定位
图11. 考据SE-hub RNA的靶基因部分参议旅途归纳
2. 功能缺失性实践
枢纽技巧:LNA敲降,dCas9-KRAB/ dCas9-LSD1
在笃定了可能与非编码区域致病突变联系的超等增强子或其靶基因之后,咱们不错通过多种分子实践来考据其功能机制。最经典且可靠的,等于功能缺失性/得回性实践。
采纳LNA longRNA GapmerRs敲降hub RNA的抒发,或通过dCas9-KRAB/ dCas9-LSD1等扼制超等增强子,通过双荧光素酶申报基因实践检测增强子活性,qPCR技巧考据下流调控基因的抒发水平。
此外,还不错使用dCas9-p300的表不雅遗传激活系统,进行功能得回性实践。
表3. 基于CRISPR/dCas9的表不雅遗传激活和扼制系统

· 案例解读 ·
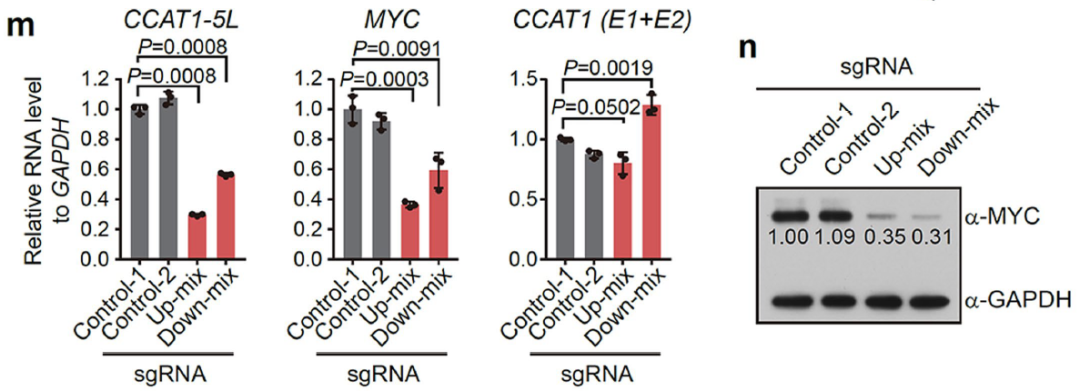
图13. 功能缺失性实践部分参议旅途归纳
3. 随意SE-hub RNA的三维结构(loop)
枢纽技巧:RADICL-seq,HiChIP-seq,Hi-C,3C-qPCR
前边RIC-seq所经营的三维结构,不错通过其他技巧得回更多的凭证,进一步考据。比如不错应用RADIBL-seq、HiChIP、Hi-C或3C-qPCR技巧,考据hub RNA与起初子RNA、eRNA在三维结构上是否存在相互战役的空间条目(比如染色指环),杀青远端(long-range)转换。
常识卡片:RADICL-seq
RADICL-seq是一种在全基因组领域内检测RNA与DNA相互作用的新设施,绘图细胞核内RNA与染色质的相互作用图谱,可应用于明白RNA分子介导的转录调控机制、构建三维基因组调控收集以及破解非编码区、增强子区突变所介导的调控失常
· 案例解读 ·
薛参议团队使用3C-qPCR进行定量染色体构象拿获实践,流露CCAT1-5L、PVT1和MYC位点之间的染色质环在CCAT1-5L缺失机显赫减少。这个数据维持了CCAT1-5L与MYC起初子RNA和eRNA相互作用以转换费力染色质环的模子,诠释超等增强子RNA与起初子RNA之间的互作可调控染色质构象。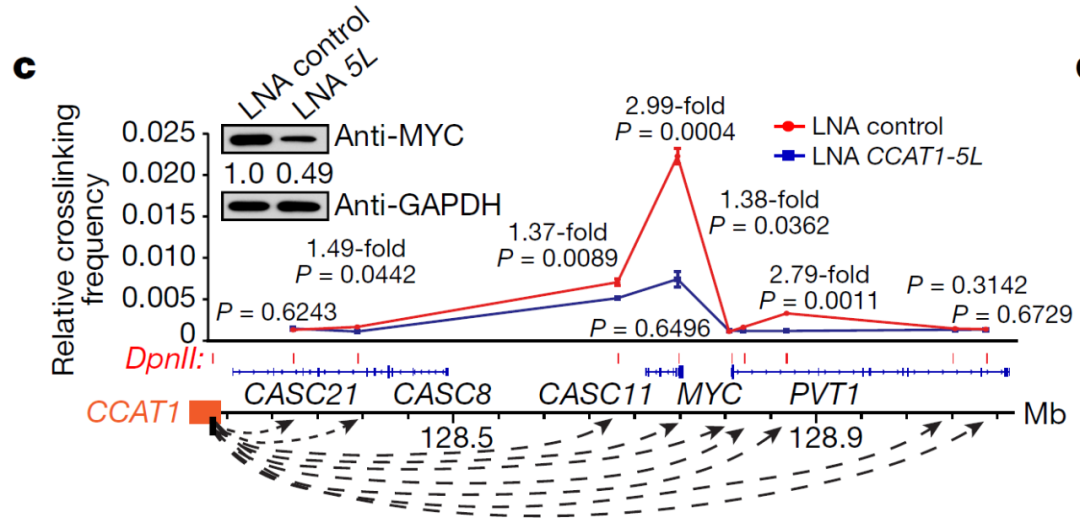
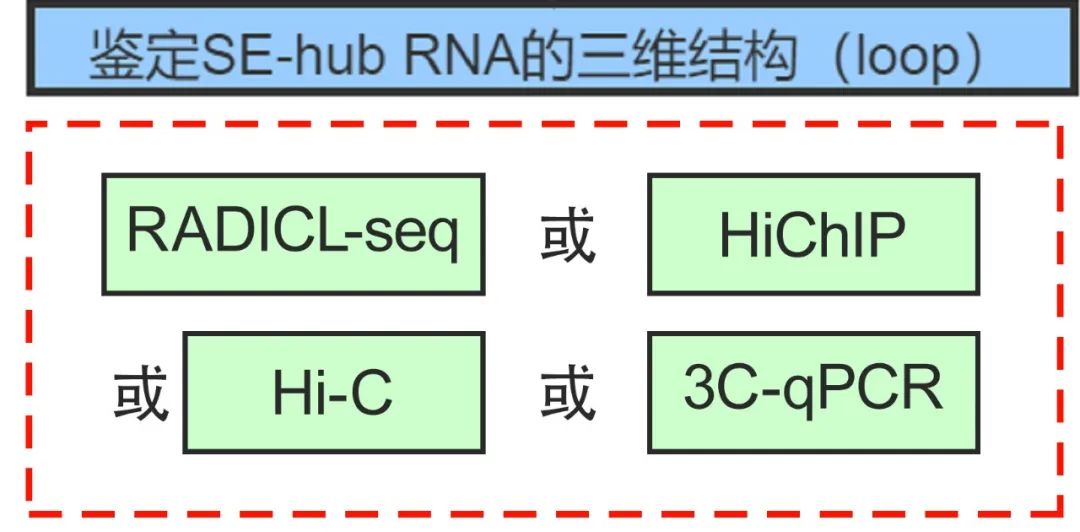
图15. SE-hub RNA三维结构随意部分参议旅途归纳
4. 寻找介导调控的RBP,并从细胞水平考据
枢纽技巧:LACE-seq,RIP-seq,CLIP-seq
东说念主类基因组编码了约1500个RNA迷惑卵白(RNA-binding protein, RBP),它们频频通过迷惑RNA分子上的特定基序或结构元件而调控细胞内各式RNA分子的加工、定位、翻译和踏实性等。在这一步,咱们不错探索SE-hub RNA参与的致病机制中,介导调控的RBP,也等于这个hub RNA上的RBP,更深切的挖掘疾病联系SE-hub RNA所依赖的调控靶基因的机制。
现在常用的随意RBP靶标的设檀越要有RIP-seq和CLIP-seq。不外这两种设施皆依赖于特异性抗体富集RBP过火所迷惑的RNA,样本所需细胞数目也多。咱们还有一个新的选定:LACE-seq。这是一项可在微量细胞中随意RBP作用靶点的新技巧,它通过线性扩增逆转录酶在RBP迷惑位点处的闭幕信号随意RBP靶标,达到单碱基分辨率的精确随意。
通过LACE-seq技巧,寻找hub RNA上的RBP;CLIP-seq进一步考据RBP与三种RNA是否存在相互作用。找到介导的RBP之后,至此,超等增强子hub RNA的基因调控机制已参议得杰出深切了。
· 案例解读 ·
薛参议团队为了明白SE-hub RNA CCAT1-5L激活MYC转录的预防分子机制,应用了ChIRP-MS、CLIP-seq、ChIP-qPCR等妙技,发现RNA迷惑卵白hnRNPK可与CCAT1-5L、MYC起初子RNA和增强子RNA PVT1相互作用,并通过coIP实考据实hnRNPK变成同型二聚体,并与RNA团聚酶II相互作用,把后者超等增强子区寄递到MYC的起初子,从而促进MYC基因的转录。
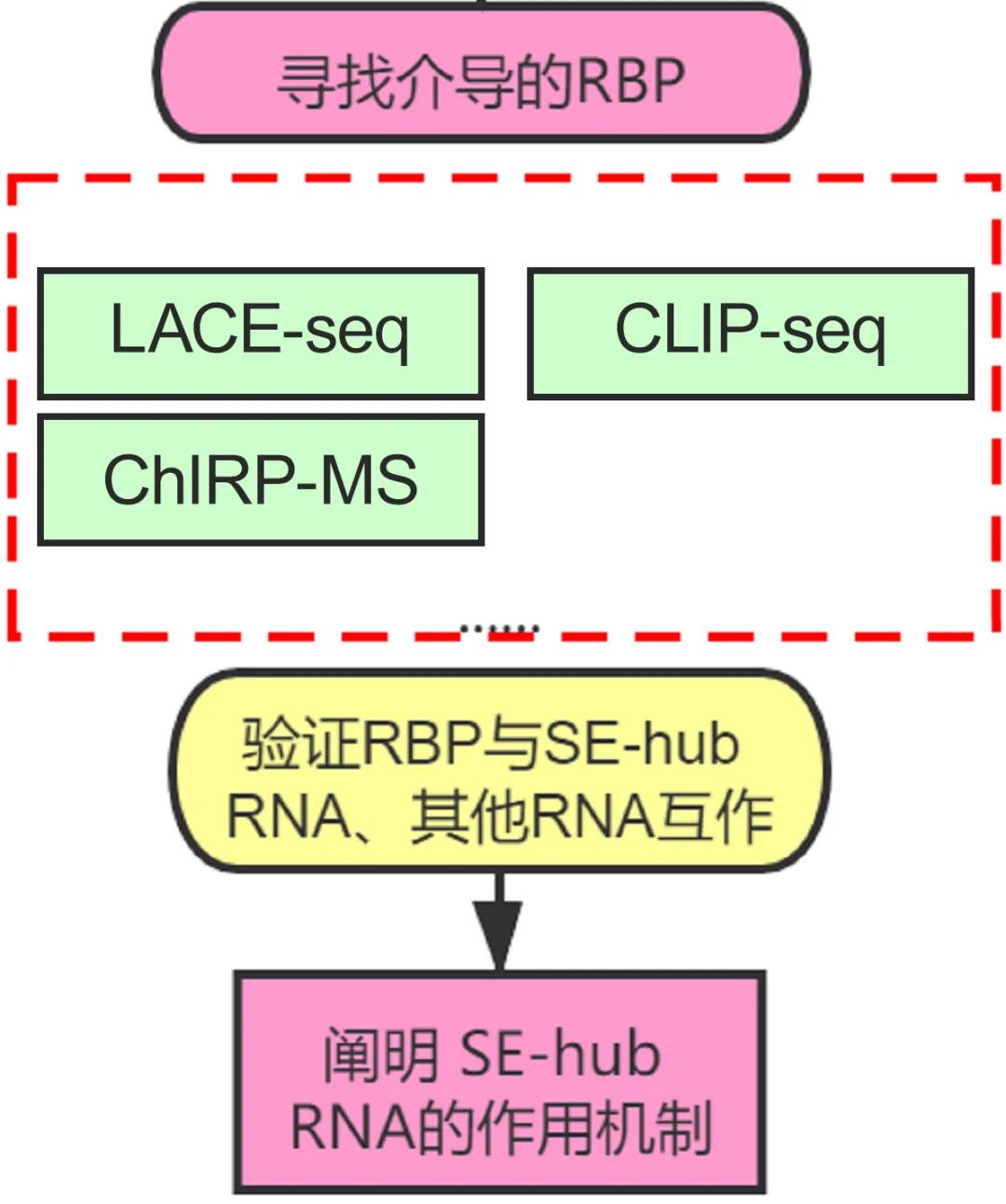
图17. 寻找介导的RBP并考据部分参议旅途归纳
写在临了
关于非编码基因的驱动作用,咱们了解的太少,未知的还有太多。现在,咱们的分析正在从外显子组逶迤为全基因组分析,意味着咱们挖掘非编码区域调控驱动事件的智力正在不停栽种。在这个领域找到新发现,为各式疾病发生发展提供更多信息,找到某种疾病发展联系的新的致病突变基因以及笃定新的标志物,是一项紧迫的任务。关联词,中枢的挑战是从比编码基因组大50倍以上非编码基因组产生的无数非功能性“乘客突变”等分离出紧迫的“驱动突变”。可幸的是,咱们领有了更多不同检测技巧,咱们有着一以贯之的坚执,为在细胞和组织水平上寻找东说念主类疾病发生发展中波及的非编码区域驱动突变事件而死力。在这里,超等增强子hub RNA动作一种中枢调控元件,将表不雅基因组和表不雅转录组跨界和谐,有望成为一个助力非编码区致病突变参议的新兴标的,让咱们拭目以俟。参考文件